Application research of Testosterone acetate
Introduction
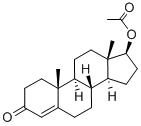
Testosterone acetate (Figure 1) is a steroid hormone synthesized and metabolized in human body. It is the main component of androgen and has a wide range of physiological functions. Testosterone acetate plays an irreplaceable role in the physiological and pathological process of human life. It not only affects the growth and development of human body and various physiological functions of the body, but also affects men's mental mood and social psychological behavior. [1-2]Testosterone is a hormone used to treat hypogonadism, breast carcinoma in women, or the vasomotor symptoms of menopause. Testosterone is a steroid sex hormone indicated to treat primary hypogonadism and hypogonadotropic hypogonadism.Testosterone antagonizes the androgen receptor to induce gene expression that causes the growth and development of masculine sex organs and secondary sexual characteristics.

Application research example
1.Testosterone fails to prevent skeletal muscle atrophy from glucocorticoids
This study was undertaken to determine whether testosterone can stimulate muscle growth as well as counteract the muscle wasting caused by excess glucocorticoids. Female rats were divided into four groups: a vehicle (1% carboxymethycellulose)-treated group, a testosterone acetate (Te)-treated group, a cortisol acetate (Co)-treated group, and a Te + Co group. Animals were injected with the steroids (Te, 40 mg/kg body wt; Co, 20 mg/kg body wt) subcutaneously for 12 consecutive days. Absolute gastrocnemius muscle weights were 8% heavier after testosterone treatment, (P less than 0.05), 20% lighter after glucocorticoid treatment, and 18% lighter after both treatments than those of vehicle-treated animals. However, total body weight loss was less in the Te + Co group than in the group receiving just Co. Specific binding of [3H]triamcinolone acetonide, a synthetic glucocorticoid, in femtomoles per milligram gastrocnemius cytosol protein, was similar between the vehicle-treated (52.9 +/- 4.7) and Te-treated (48.9 +/- 3.6) groups. Specific cytosol binding was also depleted to the same extent (to 11-12 fmol/mg protein) by either the Co or Te + Co injections. There was minimal competition by testosterone for glucocorticoid binding; however, most of the displacement was observed in the corticosteroid binder IB form of the activated triamcinolone acetonide-receptor complex on DEAE-cellulose chromatography. We conclude that Te was ineffective in preventing muscle atrophy caused by Co treatment despite its ability to induce muscle growth. The binding specificity studies lead to the conclusion that most of the testosterone and cortisol functioning were occurring primarily through separate receptor systems.[3]
2.Testosterone prevents castration-induced reduction in peripheral benzodiazepine receptors
The effect of surgical castration of adult male Sprague-Dawley rats on peripheral and central benzodiazepine (BZ) receptors was studied. Following removal of the testes, a significant decrease in the density of peripheral BZ receptors (PBR) was observed in Cowper's glands (71%; P less than 0.005) and the adrenal (31%; P less than 0.01), but not in the heart. Administration of testosterone acetate (TA) prevented castration-induced PBR depletion. Orchiectomy per se, as well as testosterone acetate administration to castrated rats, had no effect on central or peripheral BZ receptors in whole brain without the cerebellum. These results indicate the regulatory role of testosterone in the regulation of PBR in Cowper's glands and adrenal.[4]
3.Testosterone and cyproterone acetate modulate peripheral
Sixteen days of testosterone acetate (TA) treatment in male rats induced an increase in the densities of peripheral benzodiazepine receptors (PBR) in the adrenal and Cowper's glands and a decrease in PBR density in the testis. Testosterone acetate did not alter PBR density in the heart, cerebral cortex, or pituitary, or central benzodiazepine receptor (CBR) density in the cerebral cortex or hypothalamus. The antiandrogenic agent cyproterone acetate induced a decrease in PBR density in the testis, adrenal, and pituitary, but did not affect PBR density in Cowper's glands, heart, or cerebral cortex, or CBR density in the cerebral cortex or hypothalamus. In all of the above organs, affinity values did not change following the treatment with both agents. The receptoral changes may be relevant to the physiological and neurobehavioral effects of the chronic exogenous androgenic and antiandrogenic treatment.[5]
4.Detection of nandrolone, testosterone, and their esters
Nandrolone and testosterone are anabolic androgenic steroids occasionally abused by athletes. A sensitive, specific, and reproducible gas chromatography-mass spectrometry method for the quantitative determination of nandrolone, testosterone, and their esters in hair has been developed. The limits of quantitation of this method, based on 20 mg of hair, were 50 pg/mg for nandrolone and testosterone, 100 pg/mg for testosterone acetate, and 200 pg/mg for nandrolone-decanoate. Nandrolone-d3 and testosterone-d3 were used as internal standards. This method has been applied to the analysis of these compounds incorporated into rat and human hair. Male Long-Evans rats were given nandrolone decanoate 60 mg/kg intraperitoneally (i.p.) once daily for 10 days over a time period of 14 days. Two of the three rats contained nandrolone in the pigmented hair collected at day 21 at a concentration of 63 and 76 pg/mg, respectively. No drug was found in the corresponding nonpigmented hair. The rat hair samples that tested positive for nandrolone contained also nandrolone decanoate in concentrations of 0.9 and 1.2 ng/mg, respectively. In a separate experiment rats were given testosterone acetate 10 mg/kg i.p. once daily for five days. No testosterone or testosterone acetate was detected in the rat hair samples. Hair specimens were also obtained from four self-reported steroid users. The hair of two subjects were determined to be positive for testosterone in concentrations of 54 and 81 pg/mg. These data demonstrate that it is possible to detect the steroids nandrolone, testosterone, and nandrolone decanoate in hair after systemic administration.[6]
References
[1] Z. Chang, X. Zhang, L. Sui, L. Zou, Engineered microbial strain and its application in producing testosterone, Shenyang Pharmaceutical University, Peop. Rep. China . 2018, p. 12pp.
[2] Y. Chen, H. Ma, Y. Duan, X. Ma, L. Tan, J. Dong, C. Jin, R. Wei, Mycobacterium tuberculosis/Mycobacterium bovis triggered different variations in lipid composition of Bovine Alveolar Macrophages, Sci. Rep. 12(1) (2022) 13115.
[3] Capaccio JA, Kurowski TT, Czerwinski SM, Chatterton RT Jr, Hickson RC. Testosterone fails to prevent skeletal muscle atrophy from glucocorticoids. J Appl Physiol (1985). 1987;63(1):328-334. doi:10.1152/jappl.1987.63.1.328
[4] Weizman A, Amiri Z, Katz Y, Snyder SH, Gavish M. Testosterone prevents castration-induced reduction in peripheral benzodiazepine receptors in Cowper's gland and adrenal. Brain Res. 1992;572(1-2):72-75. doi:10.1016/0006-8993(92)90452-f
[5] Amiri Z, Weizman R, Katz Y, et al. Testosterone and cyproterone acetate modulate peripheral but not central benzodiazepine receptors in rats. Brain Res. 1991;553(1):155-158. doi:10.1016/0006-8993(91)90243-o
[6] Höld KM, Borges CR, Wilkins DG, Rollins DE, Joseph RE Jr. Detection of nandrolone, testosterone, and their esters in rat and human hair samples. J Anal Toxicol. 1999;23(6):416-423. doi:10.1093/jat/23.6.416
You may like
See also

Lastest Price from Testosterone acetate manufacturers

US $600.00/kg2025-11-24
- CAS:
- 1045-69-8
- Min. Order:
- 1kg
- Purity:
- 99
- Supply Ability:
- 999

US $20.00/box2025-08-01
- CAS:
- 1045-69-8
- Min. Order:
- 1box
- Purity:
- 99
- Supply Ability:
- in stock