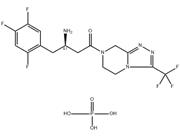
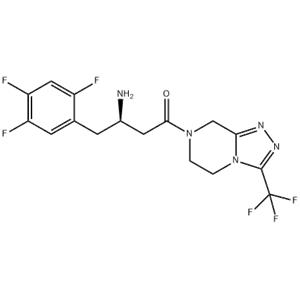
| In Vivo | In vivo, the ED50 value of sitagliptin phosphate for inhibition of plasma DPP-4 activity is calculated to be 2.3 mg/kg 7 hour postdose and 30 mg/kg 24 hour postdose in freely fed Han-Wistar rats[1]. The streptozotocin-induced type 1 diabetes mouse model exhibits elevated DPP-4 levels in the plasma that can be substantially inhibited in mice on an Sitagliptin phosphate diet. This is achieved by a positive effect on the regulation of hyperglycemia, potentially through prolongation of islet graft survival[4]. The plasma clearance and volume of distribution of Sitagliptin phosphate are higher in rats (40-48 mL/min/kg, 7-9 L/kg) than in dogs (9 mL/min/kg, 3 L/kg); and its half-life is shorter in rats,2 hours compared with 4 hours in dogs[5]. |
|---|
| Kinase Assay | DPP-4 is extracted from confluent Caco-2 cells. After 5 minutes of incubation at room temperature with lysis buffer (10 mM Tris-HCl, 150 mM NaCl, 0.04 U/mL aprotinin, 0.5% Nonidet P40, pH 8.0), cells are centrifuged at 35,000 g at 4°C for 30 minutes, and the supernatant is stored at -80°C. Assays are performed by mixing 20 μL of appropriate compound dilutions with 50 μL of the substrate for the DPP-4 enzyme, H-Ala-Pro-7-amido-4-trifluoromethylcoumarin (final concentration in the assay, 100 μM) and 30 μL of the Caco-2 cell extract (diluted 1000-fold with 100 mM Tris-HCl, 100 mM NaCl, pH 7.8). Plates are incubated at room temperature for 1 hour, and fluorescence is measured at excitation/emission wavelengths of 405/535 nm using a SpectraMax GeminiXS. Dissociation kinetics of inhibitors from the DPP-4 enzyme is determined after a 1-hour preincubation of Caco-2 cell extracts with high inhibitor concentrations (30 nM for BI 1356, 3 μM for vildagliptin). The enzymatic reaction is started by adding the substrate H-Ala-Pro-7-amido-4-trifluoromethylcoumarin after a 3000-fold dilution of the preincubation mixture with assay buffer. Under these conditions, the difference in DPP-4 activity at a certain time point in the presence or absence of an inhibitor reflects the amount of this inhibitor still bound to the DPP-4 enzyme. Maximal reaction rates (fluorescence units/seconds ×1000) at 10-minute intervals are calculated using the SoftMax software of the SpectraMax and corrected for the rate of an uninhibited reaction [(vcontrol-vinhibitor)/vcontrol]. |
|---|
| References | [1]. Thomas, L., et al. (R)-8-(3-amino-piperidin-1-yl)-7-but-2-ynyl-3-methyl-1-(4-methyl-quinazolin-2-ylmethyl)-3,7-dihydro-purine-2,6-dione (BI 1356), a novel xanthine-based dipeptidyl peptidase 4 inhibitor, has a superior potency and longer duration of action compared with other dipeptidyl peptidase-4 inhibitors. J Pharmacol Exp Ther. 2008 Apr;325(1):175-82. [2]. Kim, S.J., et al., Dipeptidyl peptidase IV inhibition with MK0431 improves islet graft survival in diabetic NOD mice partially via T-cell modulation. Diabetes, 2009. 58(3): p. 641-51. [3]. Sangle, G.V., et al., Novel biological action of the dipeptidylpeptidase-IV inhibitor, sitagliptin, as a glucagon-like peptide-1 secretagogue. Endocrinology, 2012. 153(2): p. 564-73. [4]. Kim, S.J., et al., Inhibition of dipeptidyl peptidase IV with sitagliptin (MK0431) prolongs islet graft survival in streptozotocin-induced diabetic mice. Diabetes, 2008. 57(5): p. 1331-9. [5]. Beconi, M.G., et al. Disposition of the dipeptidyl peptidase 4 inhibitor sitagliptin in rats and dogs. Drug Metab Dispos, 2007. 35(4): p. 525-32. |
|---|

 China
China


![(R)-benzyl (4-oxo-4-(3-(trifluoromethyl)-5,6-dihydro-[1,2,4]triazolo[4,3-a]pyrazin-7(8H)-yl)-1-(2,4,5-trifluorophenyl)butan-2-yl)carbamate structure](https://www.chemsrc.com/caspic/498/1242069-63-1.png)
![(R)-tert-Butyl (4-oxo-4-(3-(trifluoromethyl)-5,6-dihydro-[1,2,4]triazolo[4,3-a]pyrazin-7(8H)-yl)-1-(2,4,5-trifluorophenyl)butan-2-yl)carbamate structure](https://www.chemsrc.com/caspic/090/486460-23-5.png)
![(2Z)-4-Oxo-4-[3-(trifluoromethyl)-5,6-dihydro-[1,2,4]triazolo[4,3-a]pyrazine-7(8H)-yl]-1-(2,4,5-trifluorophenyl)but-2-en-2-amine structure](https://www.chemsrc.com/caspic/168/767340-03-4.png)
![(2Z)-4-Oxo-4-[3-(trifluoromethyl)-5,6-dihydro-[1,2,4]triazolo[4,3-a]pyrazine-7(8H)-yl]-1-(2,4,5-trifluorophenyl)butan-2-one Structure](https://www.chemsrc.com/caspic/261/764667-65-4.png) CAS#:764667-65-4
CAS#:764667-65-4![(S)-benzyl (1,4-dioxo-4-(3-(trifluoromethyl)-5,6,7,8-tetra hydro-[1,2,4]triazolo[4,3-a]pyrazin-7-yl)-1-(2,4,5-trifluorophenyl)butan-2-yl)carbamate Structure](https://www.chemsrc.com/caspic/181/1404380-73-9.png) CAS#:1404380-73-9
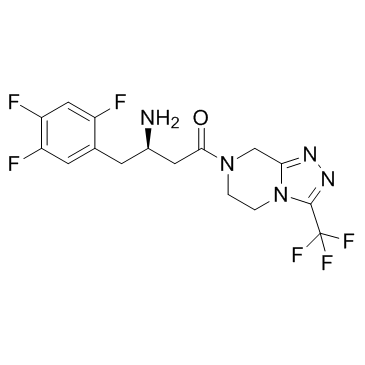
CAS#:1404380-73-9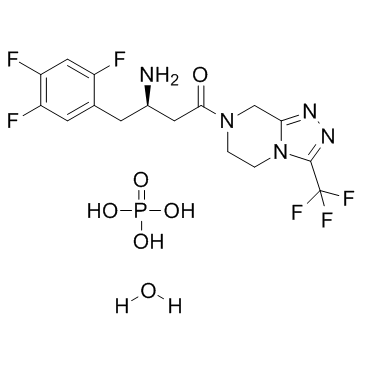 CAS#:654671-77-9
CAS#:654671-77-9