
中枢神经系统包含脑、脊髓、视网膜等组织,其生理功能的正常运转离不开精密调控的血管网络,而血脑屏障(Blood-Brain Barrier, BBB)是维系中枢内环境稳定的核心屏障结构。血脑屏障由脑微血管内皮细胞、周细胞、星形胶质细胞终足与血管基底膜协同组成,具备严格的选择性通透特性。其中内皮细胞是屏障功能的核心载体:一方面依靠高丰度紧密连接蛋白、极低的胞吞转运水平阻挡外周血液中有害物质无差别侵入中枢实质;另一方面借助内皮特异性转运蛋白与受体,完成脑部营养供给及代谢废物外排。现有大量研究证实,血脑屏障完整性受损是脑卒中、颅脑损伤、阿尔茨海默病、自闭症谱系障碍等多种神经系统疾病的共性病理特征,但中枢血管在胚胎发育阶段建立屏障特有属性、成年后持续稳定屏障功能的分子调控网络,尚未得到完整、系统的阐释。
2026年5月12日,吉林大学第一医院脑血管病学术特区畅君雷团队于《Nature Cardiovascular Research》刊发题为*CD98hc controls CNS angiogenesis and blood-brain barrier integrity through localized regulation of systemic integrin–FAK pathway*的原创研究。该工作以兼具整合素信号调控、氨基酸转运分子伴侣双重活性的跨膜糖蛋白CD98hc(SLC3A2/4F2hc)为研究对象,系统明确CD98hc在中枢血管内皮细胞中的特异性表达模式,并完整解析其调控血脑屏障发生、维持屏障稳态的分子机制。该项成果补充了中枢血管区域特化调控的全新理论依据,同时为各类中枢相关疾病的靶向干预挖掘出新的候选分子靶点。

一、CD98hc的组织内皮表达特异性特征
该研究依托多器官内皮多组学数据库结合人和小鼠组织样本染色,对比分析CD98hc在不同组织血管内皮的表达分布,明确其中枢内皮专属表达特征。 在小鼠模型中,Slc3a2编码基因mRNA在全身各器官内皮无显著表达差异,但CD98hc蛋白仅富集于脑、脊髓血管内皮,肝、肺、肾、脾、心脏等外周组织血管内皮几乎无蛋白检出,提示蛋白分布差异由翻译后调控过程介导。细胞共定位染色证实CD98hc严格定位于血管内皮细胞,不与周细胞标志物PDGFRβ、星形胶质细胞终足标志物AQP4重叠。 人类脑组织样本验证结果与小鼠表型一致,人脑皮质、海马、基底节、小脑血管内皮均高表达CD98hc,外周器官血管呈阴性。从发育时序来看,小鼠胚胎E11.5阶段脑与脊髓内皮即启动CD98hc表达,该特异性表达模式从胚胎发育持续维持至成年,提示CD98hc特异性表达是中枢血管形成屏障特性的基础前提。
二、CD98hc介导胚胎期中枢血管生成与血脑屏障构建的分子机制
为明确CD98hc生理功能,研究构建内皮特异性诱导敲除(Slc3a2 ECKO)小鼠开展胚胎层面功能验证与机制挖掘。
(一)敲除CD98hc引发特异性中枢血管畸形与屏障破裂
胚胎E13.5时期,内皮CD98hc缺失仅造成中枢血管病变,外周血管结构不受影响:小鼠大脑皮层、神经节隆起毛细血管网络稀疏、管径异常增粗、异常血管团簇大量形成;内皮细胞增殖活性下降,迁移相关丝状伪足数量、长度显著降低。至E15.5,约40%敲除胚胎发生死亡,存活个体半数伴随颅内出血。 屏障功能层面,CD98hc缺失同步破坏血脑屏障两条物质转运通路:紧密连接蛋白Claudin-5、ZO-1及抑制转胞吞关键分子Mfsd2a表达下调,介导跨胞转运的Caveolin-1、Plvap表达上调;荧光示踪剂70 kDa葡聚糖可自由渗漏进入脑实质,证明血脑屏障结构与通透功能全面受损。
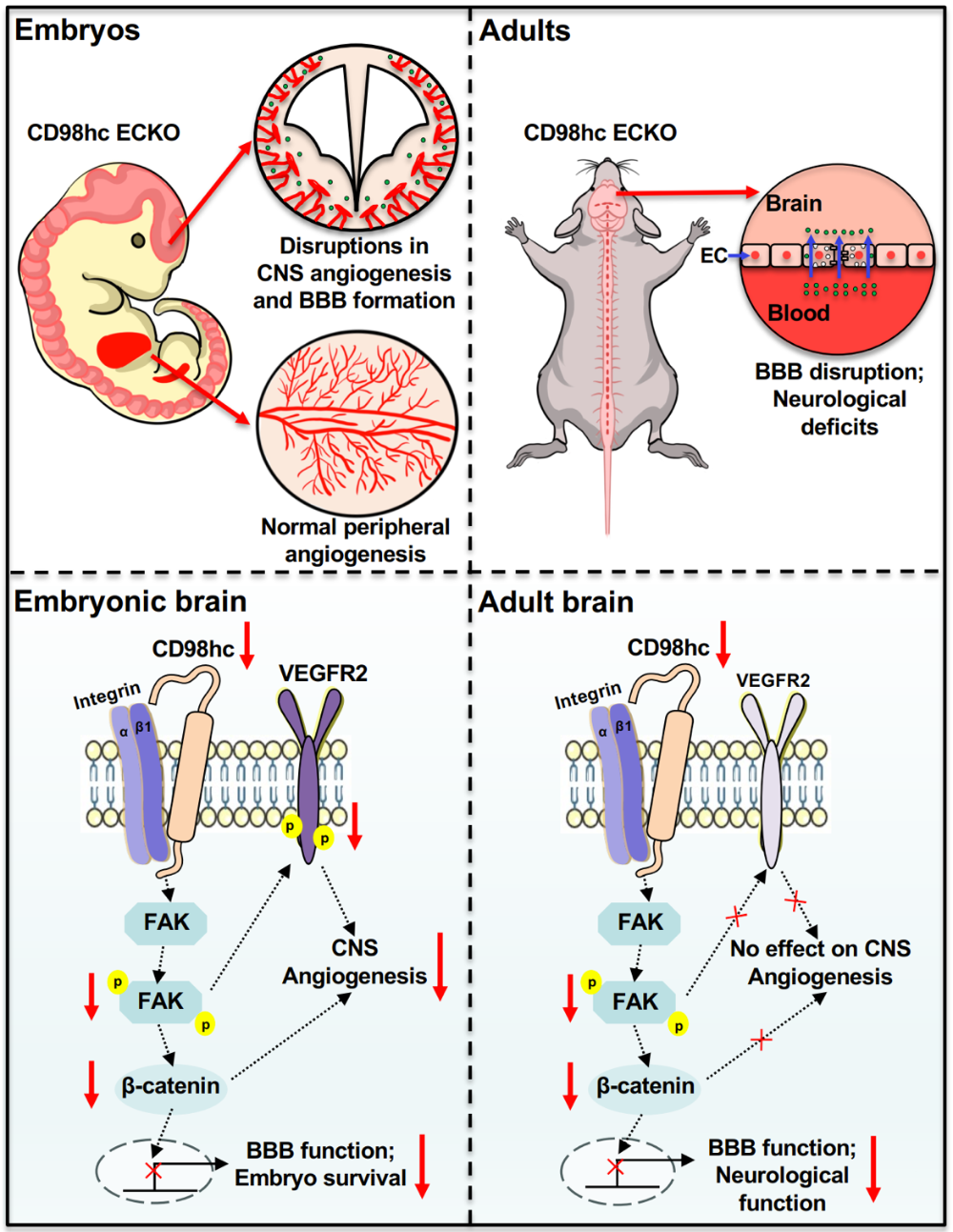
图1. CD98hc调控中枢血管生成和血脑屏障功能的作用与机制示意图
(二)Integrin-FAK为CD98hc下游核心特异性调控通路
机制实验锁定Integrin-FAK信号轴为介导CD98hc中枢血管调控功能的核心通路。体外脑血管内皮细胞敲低CD98hc不会改变Integrin β1总蛋白表达,但显著抑制FAK Y397、Y576/577位点磷酸化;免疫共沉淀结果显示CD98hc缺失会阻碍Integrin β1招募talin、vinculin、paxillin、FAK,抑制黏着斑复合体组装。表面等离子体共振试验证实CD98hc跨膜及胞内结构域可直接结合Integrin β1胞质端,KD值为615 nM,但二者不存在直接结合FAK的作用。 下游级联信号检测显示,CD98hc缺失伴随AKT、GSK3β、VEGFR2磷酸化水平下降,Wnt/β-catenin通路活性被抑制;敲除胚胎脑组织内皮pFAK、pVEGFR2及Wnt通路靶基因Lef1表达显著下调,且该通路抑制效应仅存在于中枢血管,外周血管FAK磷酸化水平无明显变化,充分体现通路调控的组织区域特异性。 2.3 FAK激动剂可逆转CD98hc缺失相关胚胎病理表型 功能挽救实验验证Integrin-FAK通路的核心作用:采用FAK特异性小分子激动剂Zn27干预敲除内皮细胞,可恢复pFAK、pVEGFR2、Wnt/β-catenin通路活性并上调紧密连接蛋白;胚胎期给予Zn27处理,能够改善敲除小鼠颅内出血、血管形态畸形、内皮Glut1表达降低等表型,恢复VEGFR2及Lef1表达,证实CD98hc缺失诱发中枢血管病变的核心诱因是FAK信号活性不足。
三、CD98hc对成年小鼠血脑屏障完整性及神经功能的调控作用
该研究进一步利用成年诱导型内皮敲除小鼠,探究CD98hc在成熟中枢血管中的生理作用。 敲除后小鼠脑血管密度、分支、总长度无明显改变,说明CD98hc不参与成熟血管结构维持,但对血脑屏障通透功能必不可少。功能检测可见443 Da小分子生物素、150 kDa大分子IgG大量从血管渗漏至脑实质;电镜下脑血管内皮紧密连接间隙增宽,介导转胞吞的小窝囊泡数量显著增多,HRP标记囊泡占比上升。 长期CD98hc缺失会继发持续性神经损伤:脑组织小胶质细胞异常活化、星形胶质细胞反应性增生,海马区NeuN阳性神经元数量减少。配套行为学检测发现模型小鼠出现多重神经功能缺陷,包括空间记忆受损、焦虑样行为增加、社交偏好与社交新奇识别能力下降。 连续两周给予Zn27干预成年敲除小鼠,可完全抑制血液大分子、小分子向脑实质渗漏,缓解胶质细胞异常活化、减少神经元丢失,同时修复物体识别、社交相关行为缺陷,再次证实FAK信号是成年阶段CD98hc调控血脑屏障功能的关键介质。
四、现有研究的科学价值与转化提示
(一)理论创新价值
1. 提出中枢血管区域特化调控新模式:CD98hc依托中枢内皮特异性表达,将全身性Integrin-FAK信号转化为中枢血管专属调控信号,协同调控VEGFR2、Wnt/β-catenin通路,分别参与胚胎血管新生与血脑屏障形成,为不同器官血管分子、功能差异化机制研究提供全新理论框架。
2. 明确CD98hc双重屏障调控功能:该分子可同时调控紧密连接组装与内皮转胞吞水平,且成年阶段对屏障功能的调控独立于血管宏观结构,是血脑屏障稳态调控的关键靶点分子。
(二)临床与转化研究提示
1. FAK激动剂具备脑病干预潜力:小分子FAK激动剂Zn27可逆转CD98hc缺失诱发的血管畸形、屏障损伤及神经功能衰退,为急性脑卒中、脑小血管病、血管性痴呆等以血脑屏障破损为基础的疾病提供全新干预思路。
2. 为CD98hc靶向脑部给药研发提出安全风险提示:已有研究将CD98hc作为介导药物跨血脑屏障递送的靶点,而该研究证实干扰CD98hc功能会直接破坏屏障完整性,造成血液成分非特异性脑内渗漏,为脑部靶向载体、抗体药物开发划定重要安全边界。
3. 搭建血脑屏障紊乱与神经精神疾病的关联证据链:已有遗传学数据显示SLC3A2突变与自闭症谱系障碍存在关联,本研究动物模型同步出现记忆、社交行为缺陷,佐证血脑屏障功能异常是部分神经精神疾病的重要中间病理环节。
五、当前研究存在的局限性与后续研究方向
从现有试验体系来看,该研究仍存在多个待深入挖掘的科学问题,也是后续该领域重点研究方向:
1. CD98hc仅在中枢内皮发生特异性蛋白富集的翻译后调控机制尚未阐明;
2. Integrin-FAK信号轴与Wnt/β-catenin通路上下游的分子连接关系有待细化完善;
3. CD98hc同时具备整合素信号调控、氨基酸转运分子伴侣两种功能,两种功能各自对血脑屏障稳态的独立贡献,需借助功能分离突变体小鼠进一步区分;
4. FAK激动剂Zn27用于脑病干预的长期药效、系统性安全性缺少完整评估数据;
5. CD98hc-Integrin-FAK通路在人脑缺血、脑出血、脑小血管病、血管性痴呆等真实临床病变中的病理作用,以及临床转化可行性,仍需大量临床样本与动物模型验证。
六、小结
该研究完整证实CD98hc是中枢血管内皮特异性标志物,依靠Integrin-FAK通路分阶段调控胚胎中枢血管发育与成年期血脑屏障完整性,其功能缺失会同步诱发血管结构畸形、屏障渗漏、神经元损伤及神经行为异常。相关成果完善了中枢血管区域特化、血脑屏障稳态维持的分子调控体系,为脑血管病、神经精神疾病机制解析与靶向药物研发提供重要实验依据;同时该研究提出的科学局限,也为领域内后续机制探索、转化医学研究提供清晰方向。
七、CD98hc检测服务哪个公司有?
LabEx 可提供专业的中枢血管与血脑屏障相关机制研究检测技术服务。平台拥有 Luminex 多因子检测、抗体芯片、蛋白磷酸化定量、共定位免疫荧光、多组学蛋白修饰分析等成熟技术,可完成血管内皮标志物定量、通路蛋白磷酸化检测、紧密连接蛋白表达分析、细胞共定位验证、炎症因子筛查等实验项目,全面适配中枢血管发育机制、血脑屏障稳态调控、脑血管疾病靶点挖掘等各类科研需求
